
达尔文的《物种起源》让全世界人民都知道了我们是从猿猴进化来的。随着研究的深入,现今考古学描绘出了人类进化的这样一幅大致图景:由于东非大裂谷的形成,非洲的古猿在进化路上分道扬镳——大裂谷以西气候变化不大,古猿进化成现今的猿类;大裂谷以东气候干旱,森林消失变成草原,一部分猿类下地生活并进化成南方古猿,他们生存于约450万-100万年前,被称为“正在形成中的人”,比如那位著名的“人类学女神”——“露西”[1]。由此,人科动物的历史正式开始[2]。
· 约200万到150万年前,南方古猿中的一支进化成能人(早期猿人)。这是最早的人属动物。
· 约170万到20万年前,能人进化成直立人(晚期猿人)并第一次走出非洲,遍布亚欧大陆,其中就包括北京猿人。
· 约20万到3万年前,早期智人阶段。这其中有从第一次走出非洲的晚期猿人独立演化而来的尼安德特人;也有第二次走出非洲并遍布亚欧的早期智人,比如许家窑人。
· 约10万到1万年前,晚期智人出现。他们第三次走出非洲,踏遍除南极洲之外的全世界每一片土地,并逐渐演化成了我们[3]。
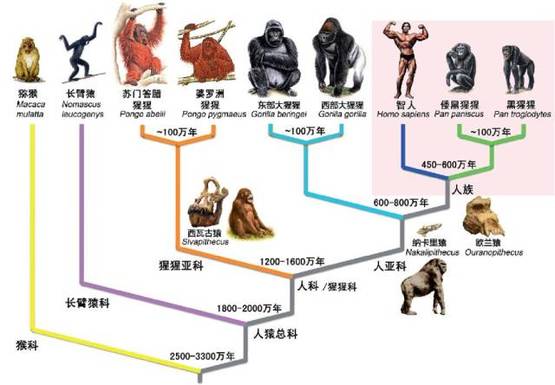 猿类进化分支谱系。图片来源:参考文献[4]
猿类进化分支谱系。图片来源:参考文献[4]
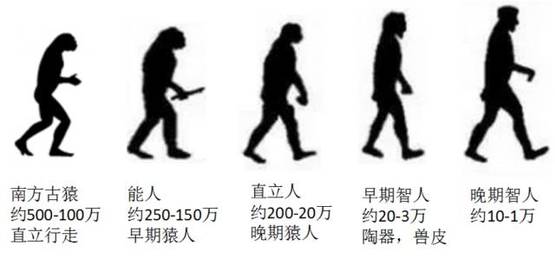 人类进化的四个主要阶段。图片来源:参考文献[10]
人类进化的四个主要阶段。图片来源:参考文献[10]
人类漫长的进化史中,近500万年的化石证据保存尚好,因此提供了较多的进化线索。不过,与浩瀚的人类进化史相比,这些证据依然不过是沧海一粟。还有很多问题需要化石证据来解答——500万年前,古猿是如何进化成类人猿的?2000万-1000万年前,我们的祖先又是什么样子?由于化石证据的缺乏,这段历史仅能靠为数不多的牙齿颌骨等化石管窥一二,在很多相关学者的眼中,这未尝不是一种遗憾。
幸好!在肯尼亚发现的一个婴儿头骨化石,弥补了这个遗憾,打破了这上百万年的空白。
 Alesi,在肯尼亚发现的已灭绝古猿的新种。这块化石的编号为KNM-NP 59050。这个新的猿种被命名为Nyanzapithecusalesi。图片来源:Fred Spoor | Copyright holder email address:meredith@leakeyfoundation.org
Alesi,在肯尼亚发现的已灭绝古猿的新种。这块化石的编号为KNM-NP 59050。这个新的猿种被命名为Nyanzapithecusalesi。图片来源:Fred Spoor | Copyright holder email address:meredith@leakeyfoundation.org
这块化石由斯图亚克布鲁克大学图纳卡纳盆地研究所的Isaiah Nengo,和美国德安扎学院领导的国际小组共同发现,编号KNM-NP 59050。发现者之一的约翰·伊库西(John Ekusi)为它起了个昵称——Alesi,在当地图尔卡纳语言中,这是祖先的意思。科学家根据Alesi臼齿的形状将它归于Nyanzapithecus属[5]。但Alesi的牙齿比同属其他物种更大,因此是一个新物种[6]。
 2014年9月在Napudet发现Alesi时,图拉卡纳盆地研究所的现场工作人员,从左到右Abdala Ekuon,John Ekusi,Isaiah Nengo,Bernard Ewoi,Akai Ekes和Cyprian Nyete。图片来源:Isaiah Nengo | Copyright holder email address: meredith@leakeyfoundation.org
2014年9月在Napudet发现Alesi时,图拉卡纳盆地研究所的现场工作人员,从左到右Abdala Ekuon,John Ekusi,Isaiah Nengo,Bernard Ewoi,Akai Ekes和Cyprian Nyete。图片来源:Isaiah Nengo | Copyright holder email address: meredith@leakeyfoundation.org
根据地质学家对化石所在岩层氩同位素测量计算,估计这块化石的年龄为1300万年。那时地球正处于中新世,即从大约二千五百万年前到五百万年前的地质时代。在这一时期,猿类出现,并遍布亚欧非三大陆,人科动物发生了广泛的进化辐射,产生了至少30个属及其中40多个被广泛认可的物种[7]。
虽然有这么多的物种,但其中只有约三分之一是从颅骨的部分遗存中知道的。而且,能保留包括脸部和腭部等重要部分的化石不超过半数[8]。在距今1700万至700万年前这一段时间,科学家尚未发现任何比较完整的颅骨化石。尤其是在1400万到1000万年前这一段时间,没有任何颅骨化石资料[9]。这大大限制了科学家对人科动物颅骨进化、形态学方面的了解。人类和现存猿类是如何分开演化的?没有新化石证据带来的线索,没人知道答案。
 考古学家的工作——迷你扫地僧。图片来源:Isaiah Nengo | Copyright holder email address: meredith@leakeyfoundation.org
考古学家的工作——迷你扫地僧。图片来源:Isaiah Nengo | Copyright holder email address: meredith@leakeyfoundation.org
而Alesi的化石,不仅是迄今为止发现的已灭绝古猿类的最完整头骨,而且还生活于1300万年前,正好处于古猿向人的远祖进化这段至关重要但缺乏考古证据的时期[5]。超级重要!
Alesi被发现的地方很久以前就被火山灰覆盖着,所以科学家猜测火山爆发可能是Alesi死亡并被埋葬的原因。

 刚刚发掘出的头骨,以及清理后头骨的真面目。图片来源:Isaiah Neng | 摄影:Christophoer Kiarie | Copyright holder email address: meredith@leakeyfoundation.org
刚刚发掘出的头骨,以及清理后头骨的真面目。图片来源:Isaiah Neng | 摄影:Christophoer Kiarie | Copyright holder email address: meredith@leakeyfoundation.org
化石显示,Alesi的牙床中全是婴儿牙齿,没有成年牙齿,因此初步推测,Alesi还只是一个婴儿。由于牙齿生长时就像树的年轮一样会留下每天的生长线,所以观察生长线即可计算其年龄。虽然在化石形成过程中牙齿已经破碎,但齿根依然存在。科学家Paul Tafforeau使用敏感度非常高的的X射线成像,观察到了尚存的齿根中的生长线,并计算其死亡时的年龄约为一年零四个月(485±40天)。但由于在人类和猿类中,将男性与女性区分开来的头骨特征在一定年龄后才会出现,因此我们并不知道Alesi的性别。

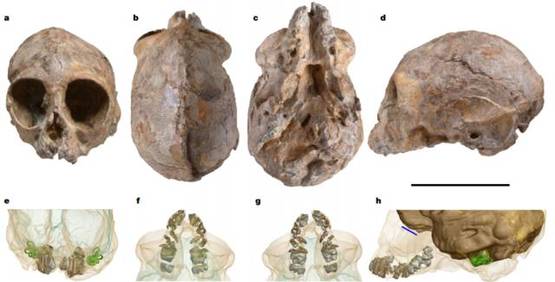 一双美丽的大眼睛,头骨多光亮。KNM-NP 59050化石的照片。a-d分别为前视图、上视图、下视图和左视图。e-h为基于X射线显微镜的三维可视化重建,视角与a-d对应,其中骨骼设置为基本透明,以显示乳牙根(浅褐色),未被破坏的永久性牙冠(灰色),骨迷路(绿色)和颅腔模型(在e-g中为蓝色部分,h中为褐色部分;由蓝线标记的嗅窝位于直线正下方)。 比例尺:5厘米。图片来源:参考文献[6]
一双美丽的大眼睛,头骨多光亮。KNM-NP 59050化石的照片。a-d分别为前视图、上视图、下视图和左视图。e-h为基于X射线显微镜的三维可视化重建,视角与a-d对应,其中骨骼设置为基本透明,以显示乳牙根(浅褐色),未被破坏的永久性牙冠(灰色),骨迷路(绿色)和颅腔模型(在e-g中为蓝色部分,h中为褐色部分;由蓝线标记的嗅窝位于直线正下方)。 比例尺:5厘米。图片来源:参考文献[6]
我们这位远古的“先祖”颜值如何呢?
科学家称,与灵长类动物相比,Alesi看起来最像是一只婴儿长臂猿。Alesi的头骨只有柠檬大小,尤其是鼻子相对偏小,与长臂猿类似。虽然科学家没有挖掘到它的身体部分,但是我们可以通过一些线索推测其运动方式。在它颅骨的内耳中具有完全发育的骨耳管,这是现代猿类的一个重要特征。长臂猿以其在树林中的快速和杂技行为而闻名,而Alesi的内耳结构表明,它可能以一种比长臂猿更慢更谨慎的方式到处走,因此Alesi可能并没有像长臂猿那样长的手臂[11]。
Alesi的发育情况怎样呢?
科学家给出了这样的进化程度分析。下图a中可以看到Alesi的颅内容积和体重的比值处于中等水平。这也反映出Alesi基本处于原猴(一种原始的灵长目动物)向人进化的中间阶段。而b中Alesi所处点斜率与人和猿类基本一致,这表明Alesi已经开始下地像猿类那样生活。
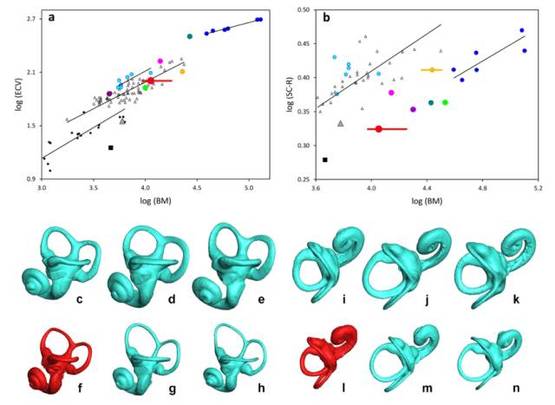 颅内容积和骨迷路。a:颅内体积(ECV,单位:毫升)和b:三个半规管的平均半径(SC-R以毫米为单位)对体重(以克计)的双变量双对数曲线。其中,蓝点为现存人类,青色点为长臂猿,小空心三角形为长尾猴属,a中小黑色方块为原猴,黑色大方块为埃及猿,a中紫色点为Pliobates,b中紫色点为Saadinius,灰色三角形为维多利亚猿,品红色点为Ekembo,橙点为山猿,绿点为Hispanopithecus,茶色点为鲁道古猿,红点为KNM-NP 59050。图中给出了现存人类、长臂猿(仅a图)、猕猴和原猴(仅a图)的压轴回归线[12-15],线段表示该属的体重值范围。c-n:左骨迷宫的左侧(c-h)和上方(i-n)视图。对应物种与图片:黑猩猩(c、i),大猩猩(d,j),苏门达腊猩猩(e,k),KNM-NP 59050(f,l),合趾猿(g,m)和灰长臂猿(h,n)。比例尺为5mm。现存物种的半规管由每种物种6到39个标本的平均形状表示。图片来源:参考文献[6]
颅内容积和骨迷路。a:颅内体积(ECV,单位:毫升)和b:三个半规管的平均半径(SC-R以毫米为单位)对体重(以克计)的双变量双对数曲线。其中,蓝点为现存人类,青色点为长臂猿,小空心三角形为长尾猴属,a中小黑色方块为原猴,黑色大方块为埃及猿,a中紫色点为Pliobates,b中紫色点为Saadinius,灰色三角形为维多利亚猿,品红色点为Ekembo,橙点为山猿,绿点为Hispanopithecus,茶色点为鲁道古猿,红点为KNM-NP 59050。图中给出了现存人类、长臂猿(仅a图)、猕猴和原猴(仅a图)的压轴回归线[12-15],线段表示该属的体重值范围。c-n:左骨迷宫的左侧(c-h)和上方(i-n)视图。对应物种与图片:黑猩猩(c、i),大猩猩(d,j),苏门达腊猩猩(e,k),KNM-NP 59050(f,l),合趾猿(g,m)和灰长臂猿(h,n)。比例尺为5mm。现存物种的半规管由每种物种6到39个标本的平均形状表示。图片来源:参考文献[6]
 Alesi的系统发育分析。未缩放(a)和缩放(b)的进化树,二者的严格一致显示出KNM-NP 59050确实属于Nyanzapithecus属,并表明了其进化状态。我们展示了十个最简约的树(a中,未缩放:1382;一致性指数= 0.289;同质指数= 0.711;保留指数= 0.597)和三个最简约的树(b中,已缩放:TL = 1274.25;一致性指数= 0.288;同质指数= 0.712 ;保留指数= 0.599)。由此可见Alesi与其他物种的关系。图片来源:参考文献[5]
Alesi的系统发育分析。未缩放(a)和缩放(b)的进化树,二者的严格一致显示出KNM-NP 59050确实属于Nyanzapithecus属,并表明了其进化状态。我们展示了十个最简约的树(a中,未缩放:1382;一致性指数= 0.289;同质指数= 0.711;保留指数= 0.597)和三个最简约的树(b中,已缩放:TL = 1274.25;一致性指数= 0.288;同质指数= 0.712 ;保留指数= 0.599)。由此可见Alesi与其他物种的关系。图片来源:参考文献[5]
发现Alesi,填补了从古猿向人进化初期的化石空白,为人类进化过程提供了宝贵的线索,在考古学和对人类起源的研究上具有特别重要的意义。现在让我们拭目以待,看科学家如何完成人类进化史的拼图吧!(编辑:明天)
参考文献:
- Kappelman J, Ketcham R A, Pearce S, et al. Perimortem fractures in Lucy suggest mortality from fall out of tall tree[J]. Nature, 2016.
- Leakdy, 贺鉴. 非洲早期的人科动物化石[J]. 科学:中文版, 1997(10):62-67.
- 刘武,寻找人类祖先的足迹:南方古猿[J],科学,1999(4):16-20
- 李辉. 遗传学对人科谱系的重构[J]. 科学:上海, 2013, 65(2):7-12.
- Benefit, B. R., Gitau, S. N., McCrossin, M. L. & Palmer, A. K. A mandible of Mabokopithecus clarki sheds new light on oreopithecid evolution. Am. J. Phys. Anthropol. 105 (Suppl. 26), 109 (1998).
- Nengo, I., Tafforeau, P., Gilbert, C.C., Fleagle, J.G.., Miller, E.R., Feibel, C., Fox, D., Feinberg, J., Pugh, K.D., Berruyer, C., Mana, S., Engle, Z. and Spoor, F. New infant cranium from the African Miocene sheds light on ape evolution. Nature 10 August 2017.
- Fleagle, J. G. Primate Adaptation and Evolution, 3rd edn (Academic, 2013).
- Begun, D. R. in Handbook of Paleoanthropology (eds Henke, W. & Tattersall, I.) 1261–1332 (Springer, 2015).
- Hill, A. & Ward, S. C. Origin of the Hominidae: the record of African large hominoid evolution between 14 My and 4 My. Yearb. Phys. Anthropol. 31, 49–83 (1988).
- 《时间-生命》丛书“早期人类”卷,F. Clark Howell著,鲁道夫·扎灵格插画,1965.
- Ryan, T. M. et al. Evolution of locomotion in Anthropoidea: the semicircular canal evidence. Proc. R. Soc. B 279, 3467–3475 (2012).
- Spoor, C. F. The Comparative Morphology and Phylogeny of The Human Bony Labyrinth. PhD thesis, Utrecht Univ. (1993).
- Spoor, F. et al. The primate semicircular canal system and locomotion. Proc. Natl Acad. Sci. USA 104, 10808–10812 (2007).
- Smith, R. J. & Jungers, W. L. Body mass in comparative primatology. J. Hum. Evol. 32, 523–559 (1997).
- Isler, K. et al. Endocranial volumes of primate species: scaling analyses using a comprehensive and reliable data set. J. Hum. Evol. 55, 967–978 (2008).